
Лаборатория Генной Истории.
R1A
Происхождение
Палеолитические охотники на мамонтов
Гаплогруппа R * возникла в Северной Азии как раз перед Последним Ледниковым Максимумом (26,500-19,000 лет до настоящего). Эта гаплогруппа была определена в 24 000-летних останках так называемого "Мальтийского мальчика» из Алтайского края, в юго-центральной Сибири (Raghavan др. 2013). Этот человек принадлежал к племени охотников на мамонтов, которые, возможно, кочевали по Сибири и части Европы в палеолите. Аутосомно это население палеолита способствовало в основном в родословной современных европейцев и выходцев из Южной Азии, эти два региона, где гаплогруппа R также, случается, наиболее распространенным в настоящее время (R1b в Западной Европе, R1a в Восточной Европе, Центральной и Южной Азии, и R2 в Южной Азии).
Гаплогруппа R1a, вероятно, отделилась от R1 * во время или вскоре после последнего ледникового максимума. О его происхождении известно немного. Некоторые думают, что она возникла на Балканах или рядом с Пакистаном и северо-западной Индией, в связи с большим генетическим разнообразием, найденного в этих регионах. Разнообразие можно объяснить другими факторами. Балканы были местом миграций из евразийских степей целых 5000 лет, каждый из которых приносил новых сортов R1a. Южная Азия была гораздо больше населенная, чем любая другая часть мира (если сравнить с Китаем) в течение менее чем 10 000 лет, а большее население приносит большее генетическое разнообразие. Наиболее вероятным местом происхождения R1a является Центральная Азия или Юг России / Сибирь.
Оттуда R1a мог мигрировать непосредственно к Восточной Европе (европейской части России, Украина, Белоруссия), или на юг через Центральную Азию и Иран. В последнюю очередь R1a пересек Кавказ в эпоху неолита, наряду R1b колонизировал понтийско-каспийскую степь. Из-за отсутствия древней Y-ДНК из этих регионов лучшее свидетельство поддержки позднего палеолита миграции в Иран является наличие очень старых субкладов из R1a ( M420) в регионе, особенно в горах Загрос. Однако эти образцы составляют лишь малую часть всех R1a в регионе и с таким же успехом представляют потомков восточноевропейских охотников-собирателей, которые отделились от других племен R1a и ушли с Северного Кавказа между 20 000 и 8000 лет назад. Логика в том, что наиболее известные исторические миграции в Евразии проходил с севера на юг, поскольку люди искали теплые края. Единственное исключение произошло во время Голоцена разогрева климата, что соответствует неолиту колонизации Европы с Ближнего Востока. Третья возможность заключается в том, что R1a племена разделены на две части по всему Казахстану в позднем палеолите, с одной группой в Восточной Европе, в то время как другая группа двинулась на юг в Иран.
R1a пришли в Европу с неолитическими фермерами?
Некоторые люди предположили, что R1a была одной из линий неолитических фермеров, и вошла бы в Европу через Анатолию, затем распространилась по всех Балканах в сторону Центральной Европы, и только потом в страны Восточной Европы. Есть много вопросов по этому сценарию. Во-первых, 99% современного R1a происходит с R1a1a (M417), субклад, что явно расширился из бронзовой эпохи, а не с раннего неолита. Его филогения также указывает на Восточное европейское происхождение. Во-вторых, большая часть R1a в Ближнем Востоке является глубоким субкладом филиала R1a-Z93, который возник в России (см ниже). Они не могут быть предком Балканского или Центрально-Европейского R1a. В-третьих, существует очень сильная корреляция между аутосомной примесью Северо-Восточной Европы и населения R1a, и этот компонент отсутствует в геноме всех европейских неолитических фермеров протестированных на сегодняшний день - даже от Отзи, который был фермером энеолита. Это смешение также отсутствует у современных сардинцев, которые в основном происходят от фермеров неолита. Это неопровержимое доказательств того, что R1a не пришел в Европу от неолитических фермеров.
Бронзовый век Прото-индоевропейцы
Предполагается, что R1a были доминирующей гаплогруппой среди северных и восточных прото-индо-европейских носителей языка, что вовлекло их в индо-иранские, фракийские, балтийские и славянские ветви. Прото-индоевропейцы возникли в ямной культуре (3300-2500 до н.э.). Их резкое расширение стало возможным благодаря скорейшему освоению бронзового оружия и одомашниванию лошади в евразийских степях (ок 4000-3500 до н.э.). Южная степная культура предположительно унаследовала преимущественно R1b (M269 и M73), а в северной лесостепной культуре существенно доминировал R1a. Первая экспансия лесостепных людей произошла от культуры шнуровой керамики (см германский филиал ниже). Миграция R1b людей в Центральной и Западной Европе оставил вакуум для R1a людей в южной степи вокруг времени катакомбной культуры (2800-2200 до н.э.).Лесостепной происхождение этой культуры очевидна от внедрения проводной керамики и обильным использованием полированных боевые топоры, двух выдающихся особенностей культуры шнуровой керамики. Это также, вероятно, когда процесс satemisation из индоевропейских языков началось с балто-славянских и языковые группы индоиранских принадлежат к той же сатем изоглосса и оба, кажется, превратилась из катакомбной культуры.
Древняя ДНК-тестирование подтвердило присутствие гаплогруппы R1a1a в пробах, взятых в культуре шнуровой керамики в Германии (2600 г. до н.э.), от тохарских мумий (2000 до н.э.) в Северо-Западном Китае, от Курганской захоронений (около 1600 г. до н.э.) из андроновской культуры на юге России и южной Сибири, а также из различных Железный возраста сайтов из России, Сибири, Монголии и Центральной Азии.

В настоящее время, высокие частоты R1a найдены в Польше (57,5% населения), Украина (40 до 65%), европейской части России (от 45 до 65%), Беларуси (51%), Словакии (42%), Латвии (40 %), Литва (38%), Чехии (34%), Венгрия (32%), Норвегия (27%), Австрия (26%), Хорватия (24%), на северо-востоке Германии (24%) Швеция (19%) и Румынии (18%).
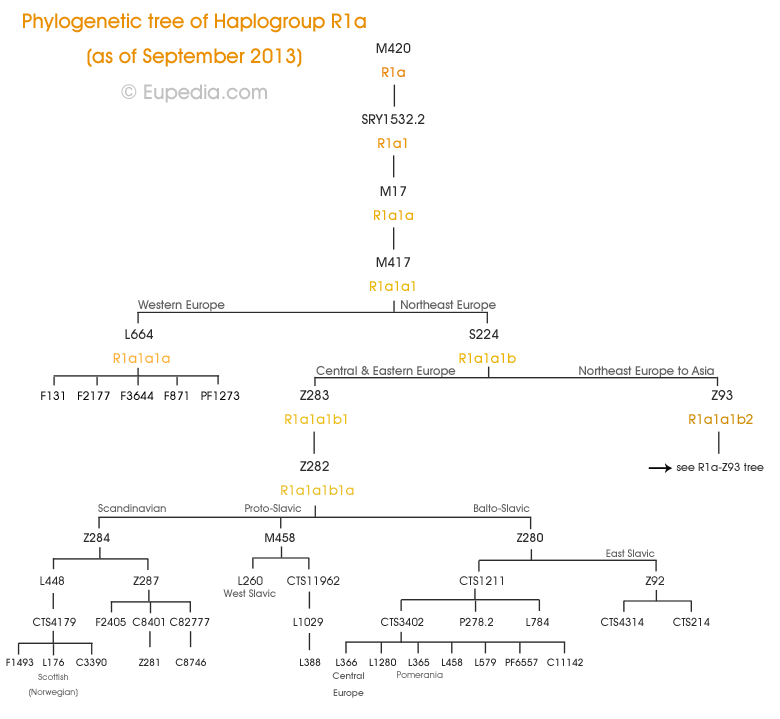
99% R1a люди принадлежат к субкладов из R1a1a1 (R1a-M417), которая делится в следующих субкладов:
R1a-L664 по существу Северо-Западный Европейский, нашел главным образом в Западной Германии, стран Бенилюкса и на Британских островах.
R1a-Z645 составляет кучу R1a лиц от Центральной Европы до Южной Азии.
R1a-Z283 является основным Центральный и Восточный европейское отделение.
R1a-Z284 является скандинавской субклад с эпицентром в Норвегии. Установлено, также в местах, колонизированных норвежских викингов, как некоторых частях Шотландии, Англии и Ирландии. Несколько субклады были определены, в том числе L448, L176.1, Z287 / Z288, Z66 и Z281, о котором мало что известно на данный момент.
R1a-M458, в первую очередь Славянский субклад, с максимальными частотами в Польше, Чехии, Словакии, но также довольно распространены в юго-восточной Украине и северо-западе России.
его субклад R1a-L260 явно западнославянских, с пиком частоты в Польше, Чехии и Словакии, и излучающих на более низких частотах в Восточной Германии, Восточной Австрии, Словении и Венгрии.
R1a-Z280 также балто-славянская маркер, найти во всем Центральной и Восточной Европы (за исключением Балкан), с западной границы работает с востока на юго-западе Германии и в Северо-Восточной Италии. Это может быть разделена на множество кластеров: восточнославянских, прибалтийских, поморских, польских, Карпат, Восточно-альпийских, чехословацких, и так далее.
его субклад R1a-L365 является Поморское кластер найдено также на юге Польши.
R1a-Z93 является основным Азии филиал R1a. Он находится в Центральной Азии, Южной Азии и Юго-Западной Азии (в том числе среди ашкеназских евреев). R1a-Z93 является маркером исторических народов, таких как индоариев, персов, Мидии, Митанни, или татар, и проникнут генофонд арабов и евреев.
его субклад R1a-M434 составляет небольшой процент населения Пакистана. Следы были также найдены в Омане.
Много Западной и Северной Европы R1a, что является отрицательным для маркера Z284 попадает под корень R1a1a1 * (M417), или даже в более старых R1a1a (M17) и R1a1 (SRY10831.2). Бывший произошли от старых известных расширения R1a из Лесостепи, на культуры шнуровой керамики (см ниже), который предшествует всем вышеперечисленным субклады. В настоящее время нет субклад не был определен общий SNP. Тем не менее, Клёсов и соавт. (2009) обнаружили, что значительная часть R1a в Северо-Западной Европе, в частности в Норвегии, Англии, Ирландии и Исландии, имели значение повтора 10 (вместо 12) на STR-маркера DYS388. Среди них, некоторые люди были идентифицированы как несущий мутацию L664. Происхождение старых субкладов (M17 и SRY10831.2) до сих пор неясно (возможно, мезолита охотники-собиратели, бродящие вокруг Европы).
История R1a
Германский филиал
Первый крупный расширение R1a состоялась с запад распространения шнуровой керамики (или боевой топор) культуры (2800-1800 до н.э.) из северной лесостепи на родине ямной. Это была первая волна из R1a в Европе, тот, который принес Z283 субклад к Германии и Нидерландах, и Z284 в Скандинавию. В шнуровой керамики R1a люди бы смешивается с предварительно германской I1 и I2 аборигенов, что привело к первой индоевропейской культуры в Германии и Скандинавии, хотя, что культура не может считаться Прото-германский - это было просто Прото-индо-Европейский на этом этапе, а может быть, или прото-балто-славянская.
Германские языки, вероятно, не появится до Северных бронзового века (1800-500 до н.э.). Прото-германский язык, вероятно, разработаны в виде смеси двух ветвей индоевропейских языков, а именно Прото-балто-славянского языка культуры шнуровой-Ware (R1a-Z283) и последующего прихода Прото-итало-кельто-германских народов из культуры Únětice (R1b-L11). Это подтверждается тем, что германский народ являются гибридом R1a-R1b, что эти два гаплогруппы пришли через отдельных маршрутов в разное время, и, что Прото-германский язык является самым близким к Прото-итало-кельтских, но и разделяет сходство с прото-Славянский.
R1b ветвь индоевропейцев, как полагают, возникла в южной культуры ямной (северных берегах Черного моря). Это было первым, чтобы перейти из степей в Европу, вторгаясь в дельте Дуная вокруг 4200 до н.э., затем вносит свой путь вокруг Балкан и венгерскую равнину в 4-м тысячелетии до н. Вполне вероятно, что меньшинство R1a людей сопровождается эту миграцию R1b. Эти R1a мужчины бы принадлежал к L664 субклада, первый раскол от ядра ямной. Эти ранние степные завоеватели не были однородной группой, но кластер племен. Возможно, что R1a-L664 люди были один или несколько отдельных племен самостоятельно, или, что они смешиваются с некоторыми линий R1b, в частности R1b-U106, которые станут основными германской линии преемственности много веков спустя.R1b-R1a контингент перешел Дунай в Паннонского равнине около 2800 г. до н.э., был положен конец местный Bell Стакан (около 2200 г. до н.э.) и шнуровой керамики (с. 2400 г. до н.э.) культур в Центральной Европе, а также создать культуру Únětice (2300-1600 до н.э.) вокруг Богемии и восточной Германии. Únětice можно рассматривать как источник будущих германских, кельтских и италийских культур, и связано с L11 субклада из R1b.
Покойный Únětice культура расширена до Скандинавии, основав Nordic бронзового века. R1a-L664 и R1b (L11 и U106) предположительно достигли Скандинавии в это время. Люди из стран Северной Европы бронзового века, вероятно, говорил Прото-германский язык, который на протяжении более тысячи лет приобретенный словарный запас от коренного языка шнуровой керамики, само по себе смесь прото-балто-славянских и не-IE предварительно германской. Первый подлинный германский язык, по оценкам лингвистов, пришли в существование вокруг (или после) 500 г. до н.э., как Nordic бронзы подошел к концу, уступая место Pre-римской железного века. Уникальность некоторых германских пунктов лексики в заимствования из родных предварительно индоевропейских языков (германская теория субстрат). Кельтский язык сам по себе, как известно, заимствовано из афро-азиатских языков, на которых говорят ближневосточных иммигрантов в Центральной Европе. Тот факт, что современная Скандинавия состоит из примерно 40% от I1, 20% R1a и 40% R1b укрепляет идею, что германский этнос и язык приобрел три-гибрид характер по железному веку.
Балтийский филиал
Балтийский филиал, как полагают, произошли от культуры фатьяновской (3200-2300 до н.э.), на северо-востоке продлении культуры шнуровой керамики. Ранние кочевники бронзового века R1a из северных степей и лесостепей бы смешивают с коренными уральская-говорящих жителей (N1c1 линий) региона. Это подтверждается сильным присутствием обоих R1a и N1c1 гаплогрупп от южной Финляндии в Литве и прилегающей части России.
Славянский филиал
Истоки славян вернуться к Circa 3500 до н.э. с северной ямной культуры. В M412 и Z280 клоны распространился по всему Польши, Беларуси, Украины и западной части России, и будет формировать ядро праславянского культуры. Высокая распространенность R1a в странах Балто-славянские на сегодняшний день является не только в связи с расширением шнуровой керамики, но и длинного ряда более поздних миграций из России, последний из которых проходил с 5-го по 1-е века н.э.. Славянский филиал дифференцированы себя, когда культура шнуровой керамики поглотили культуру Кукутень-трипольской (5200-2600 до н.э.) из Западной Украины и северо-востоке Румынии, которая, кажется, были составлены в основном из I2a1b (M423) линий, выведенный непосредственно из палеолитических европейцев, с небольшой примесью ближневосточных иммигрантов (особенно E1b1b, G2a, J и T). Так возник гибридный шаровидных амфор культуры (3400-2800 г. до н.э.) в том, что теперь Украине, Беларуси и Польши. Это, безусловно, в этот период, что I2a2, E-V13 и T распространение (наряду с R1a) вокруг Польши, Беларуси и западной части России, объясняя, почему восточные и северные славяне (и литовцы) имеют от 10 до 20% I2a1b линий и около 10 % от ближневосточных линий (18% для украинцев). Спустя всего несколько столетий, это гибридизованной культура исчезла в доминирующую шнуровой керамики (2800-1800 до н.э.) и катакомбной (2800-1800 до н.э.) культур.
Последовал период шнуровой керамики в степях по культуре Srubna (1800-1200 до н.э.), и по Польше культурой Trzciniec (1700-1200 до н.э.). Последним важным Славянский миграция, как полагают, произошло в 6-м веке нашей эры, из Украины в Польшу, Чешскую Республику и Словакию, заполняя вакуум, оставленный восточных германских племен, вторгшихся в Римскую империю.
Исторически сложилось так, никакая другая часть Европы не была захвачена большее количество раз по степных народов, чем на Балканах. Хронологически первые R1a захватчики пришли с запад расширения культуры ямной (от 4200 до н.э.), череды степных миграций, которая длилась около 2000 лет. Потом фракийцы (1500 до н.э.), после чего иллирийцев (около 1200 г. до н.э.), гуннов и аланов (400 CE), аварцы, булгар и сербы (все вокруг 600 CE), и мадьяры (900 CE ), среди прочих. Эти народы, происходящих из различных частей евразийских степей, в любом месте между Восточной Европе и Центральной Азии, и именно поэтому такая высокая STR разнообразие находится в Балканском R1a в настоящее время. Это пока не представляется возможным определить этническое происхождение для каждой разновидности R1a, помимо того, что ни о какой R1a связано с племенами из Евразийской степи в одной точке в истории.
Индо-иранская ветвь
Прото-индо-иранские колонки, люди, которые позже назвали себя «арийцами» в Ригведе и Авесте, возникла в культуре Синташтинская-Петровка (2100-1750 до н.э.), в долинах Тобола и Ишима, к востоку от Уральских гор . Он был основан скотоводческих кочевников с культурой абашевским (2500-1900 до н.э.), начиная от верхней Дон-Волга до Уральских гор, и культуры Полтавка (2700-2100 до н.э.), проходящую от нижней Дон-Волга в Каспийское море депрессия.
Культура Синташтинская-Петровка, связанные с R1a-Z93 и его субкладов, был первым бронзы продвижение индоевропейцев к западу от Урала, открывая путь к обширных равнинах и пустынях Средней Азии к металлическим богатых Алтайских гор . Арийцы быстро расширилась за всей Центральной Азии, с берегов Каспия до юга Сибири и Тянь-Шаня, через торгов, сезонных миграций стад, и грабежи рейдов.
Конные боевые колесницы, кажется, были изобретены синташтинских людей вокруг 2100 до н.э., и быстро распространился на горной области Бактрия-Маргианы (современной границе Туркменистана, Узбекистана, Таджикистана и Афганистана). Медь добывался интенсивно на Урале, и прото-индо-иранцы от синташтинской-Петровке были экспортировать ее в огромных количествах на Ближний Восток. Они, кажется, были привлечены природных ресурсов долины Зеравшан для Петровка медно-добывающей колонии была создана в тугай вокруг 1900 до н.э., и олово экстрагировали вскоре на Карнаб и Mushiston. Олово было особенно ценным ресурсом в конце бронзового века, когда оружие изготавливались из медно-оловянного сплава, сильнее, чем более примитивной мышьяком бронзы. В 1700 г. до н.э., индо-иранцы расширена до нижней Амударьи долине и поселился в орошение сельскохозяйственных общин (Tazabagyab культуры). По 1600 г. до н.э., старые укрепленные города Маргиана-Бактрии были заброшены, затоплены северных степных мигрантов. Группа культур Центральной Азии под Индо-иранского влияния известен как андроновской горизонте, и продолжался до 800 г. до н.э..
Индо-иранские миграции продвинулись дальше на юг через Гиндукуш. По 1700 г. до н.э., конные скотоводы проникли в Белуджистане (юго-запад Пакистана).Долины Инда поддался около 1500 г. до н.э., и северные и центральные части Индийского субконтинента были переданы 500 г. до н.э.. Запад миграции привело Старый индийских санскритских колонки верхом боевые колесницы в Ассирию, где они стали правителями Митанни примерно с 1500 г. до н.э.. Мидии, парфяне и персы, все иранские докладчики из андроновской культуры, переехал в Иранском нагорье от 800 до нашей эры. Те, что остались в Центральной Азии помнят историю как скифов, в то время как потомки ямной, которые остались в понтийского-Каспийского степи стал известен как сарматов к древним грекам и римлянам.
Индо-иранские миграции привели к высоким частотам R1a в южной Центральной Азии, Ирана и Индии. Наибольшая частота R1a (около 65%) достигается в кластере вокруг Кыргызстана, Таджикистана и Северного Афганистана. В Индии и Пакистане, R1a составляет от 15 до 50% населения, в зависимости от региона, этнической группы и касты. R1a, как правило, сильнее Северо-Западный субконтинента, и слабым в дравидийских-говорящей Юга (Тамил Наду, Керала, Карнатака, Андхра-Прадеш) и из Бенгалии на восток. Более 70% брахманов (высшая каста в Hindusim) принадлежат R1a1, в связи с эффектом основателя.
Материнских линий в Южной Азии, однако, в подавляющем большинстве предварительно индоевропейской. Например, в Индии более 75% «родных» мтДНК M и R линий и 10%-Восточной Азии линий. В остаточной 15% гаплогрупп, примерно половина из них из ближневосточного происхождения. Только около 7 или 8% может быть "русской" (Причерноморье-Каспийского степной) происхождения, в основном в виде гаплогруппы U2 и W (хотя происхождение U2 по-прежнему обсуждается). Европейский мтДНК происхождений встречаются гораздо чаще, в Центральной Азии, хотя, и даже в Афганистане и на севере Пакистана. Это говорит о том, что индоевропейская вторжение в Индии был проведен в основном мужчинами через войны, и первым крупным урегулирование женщин было в северном Пакистане, западная Индия (Пенджаб Гуджарат) и северной Индии (штат Уттар-Прадеш), где гаплогруппы U2 и W являются наиболее распространенными.
Мумии Тарим
В 1934 шведский археолог Фольке Бергман обнаружили около 200 мумий светловолосых кавказских народов в Таримской впадины в Северо-Западном Китае (в регионе, известном как Синьцзян, Восточном Туркестане или Uyghurstan). Самый старый из этих мумий датируются 2000 г. до н.э., и все 7 мужчин остается проходят проверку Li и соавт. (2010), были положительными для мутаций R1a1. Современные жители бассейна реки Тарим, уйгуров, принадлежат как к этому R1b-M73 субклада (около 20%) и R1a1 (около 30%).
Первая теория о происхождении мумий Тарим является то, что группа начала всадников из культуры имени И. Е. Репина (3700-3300 до н.э.) мигрировали из Дон-Поволжья в Горном Алтае, основав культуру Афанасьево (с. 3600-2400 до н.э.), откуда они двинулись на юг, чтобы бассейна реки Тарим. Другая возможность состоит в том, что Таримские мумии спуститься с прото-индо-иранского народа (см выше), которые расширенных всей Центральной Азии около 2000 года до нашей эры из культуры Синташтинская-Петровка. Ответвление бы пересек горы Тянь-Шаня, в конечном итоге в Таримской впадины. Эта теория имеет то преимущество, соответствующие датировку мумий Тарим. В любом случае, большинство из мумий, протестированных на мтДНК принадлежали к монголоидной гаплогруппы C4, и лишь немногие европейские или ближневосточные гаплогрупп (H, K и R).
Существует некоторое противоречие в отношении возможной связи между мумий Тарим и тохарских языках, Centum ветви индо-европейской семьи, которые были сказаны в Таримской впадины с 3 по 9-й века н.э.. Легко предположить, что Таримские мумии были прото-тохарийские динамики в связи с соответствующем месте и индоевропейской связи. Тем не менее, Таримские мумии предшествовали появление тохарского по более двух тысячелетий, и тохарский это язык Centum, что не может быть сошел с прото-индо-иранской ветви сатем. Другие отрасли CENTUM время все они связаны с гаплогруппы R1b, и тохарский быть единственным восточной Centum язык, это возможно, что тохарийские колонки вместо связан с Центральной Азии R1b1b1 (M73) субклада, также найденного среди современных уйгуров, населяющих бассейн Тарима.
Тюркские динамики и R1a
Современные жители Центральной Азии, из Синьцзяна в Турцию и от Волги до Гиндукуша, говорят в подавляющем большинстве тюркских языков. Это может быть удивительно, так как это соответствует области, где индоиранских к индо-европейских спикеров расширенной, культура бронзового века андроновской, и скифская территория Утюг-Возраст. Итак, почему это, что индоевропейские языки выживает только в славянской России или в южной части Центральной Азии, в таких местах, как Таджикистан, Афганистан или некоторых частях Туркменистана? Почему не уйгуры, узбеки, казахи и кыргызы или современные понтийского-каспийские степи люди (крымские татары, ногайцы, башкиры и чуваши) говорят индоевропейские наречия? Генетически эти люди несут индоевропейский R1a, и в меньшей степени также R1b, линий. Объясняется это тем, тюркские языки заменены иранские языки Центральной Азии между 4-м и 11-м веке нашей эры.
Пратюркский возникла в Монголии и Южной Сибири с такими кочевых племен как хунну. Он принадлежит к алтайской языковой семье, как монгольский и маньчжурский (некоторые также корейский и японский, хотя они разделяют очень мало лексику общего). Это неизвестно, когда Пратюркский впервые появились, но его распространение началось с гуннских миграций на запад через евразийской степи и всю дорогу в Европу, только остановился на границах Римской империи.
Гунны были потомками хунну. Древние ДНК-тесты показали, что хунну были уже гибридной Евразии людей 2000 лет назад, со смешанным Европы и Северо-Восточной Азии Y-ДНК и мтДНК. Современные жители родины хунну имеют примерно 90% монгольских линий против 10% европейских.Старая идентифицированная наличие европейской мтДНК вокруг Монголии и Байкала датируется более 6000 лет назад.
Похоже, что тюркский быстро заменить скифские и другие иранские диалекты всей Центральной Азии. Другие перелетные волны принесли более тюркские колонки к Восточной и Центральной Европы, как хазар, аваров, булгар и турками (=> см 5000 лет миграций из евразийских степей в Европу). Все они были на самом деле кочевников Центральной Азии, принявших тюркский язык, но было мало, если любой монгольской крови. Тюркские нашествия поэтому больший вклад диффузии индоевропейских линий (особенно R1a1), чем азиатские Востока.
Тюркские языки не сохранились в Европе за пределами понтийского-Каспийского степи. Болгарский язык, несмотря на то, назван в честь тюркского племени, на самом деле является Славянский язык с мягким тюркского влияния. Венгерский, иногда ошибочно принимают за наследника гуннского потому его имени, на самом деле язык уральские (Magyar). в Десятки тюркских языков в современном мире имеют высокую степень взаимного разборчивости в связи с их довольно недавнего общего происхождения и кочевого характера его носителей (до недавнего времени). Две основные ветви огузы и Oghur можно рассматривать как двух языках о качестве далекой, как испанский и итальянский, и языков в каждой отрасли, как региональных диалектов испанского и итальянского.
Греческий филиал
Мало что известно о прибытии Прото-греческих ораторов от степей.Микенской культуры началось около 1650 г. до н.э. и явно импортированы степной культуры. Тесная взаимосвязь между микенской и прото-индо-иранских языков предполагают, что они расстались довольно поздно, в период между 2500 и 2000 до н.э.. Археологически, микенские колесницы, наконечники копий, кинжалы и другие бронзовые предметы показывают поразительное сходство с культурой сейминско-турбинского (с. 1900-1600 до н.э.) из северных российских лесостепи, известные для большой подвижности его кочевых воинов (SEIMA-турбинского сайты были найдены как далеко, как Монголия). Поэтому вполне вероятно, что микенской спустился из России в Греции между 1900 и 1650 до н.э., где они смешались с местными жителями, чтобы создать новый уникальный греческую культуру.
Население R1 распространяться гены для светлой кожи, светлыми волосами и рыжими волосами
Существует в настоящее время убедительные доказательства, что оба R1a и R1b люди внесли свой вклад диффузии A111T мутации SLC24A5, что объясняет apporximately 35% от разницы кожи тона между европейцами и африканцами, и большинство вариаций в пределах Южной Азии. Характер распределения в A111T аллеля (rs1426654) матчей практически идеально распространение индоевропейской R1a и R1b линий по всей Европе, на Ближнем Востоке, Центральной Азии и Южной Азии. Мутация, вероятно, передается в раннем неолите на другие группы населения Ближнего Востока, которая объясняет, почему неолитические земледельцы в Европе уже нес A111T аллель (например Келлер 2012 стр.4, Лазаридис 2014 доп. 7), хотя при более низкой частоте, чем современных европейцев и южные страны Центральной Азии.
Свет аллель кожа также нашли в диапазоне от 15 до 30% в различных этнических групп в северной Африке южнее Сахары, в основном в странах Сахеля и саванн зон, населенных племенами R1b-V88 скотоводов, как фульбе и хауса. Это предполагало бы, что A111T аллель уже присутствует среди всех R1b людей перед Pre-Pottery неолита раскола между V88 и P297. Популяции R1a иметь столь высокую частоту этого аллеля в популяциях R1b. С другой стороны, мутация A111T отсутствовал на 24000-летней R * образца из Сибири, и отсутствует в большинстве современных популяций R2 в Юго-Восточной Индии и Юго-Восточной Азии. Следовательно, он может быть с уверенностью предположить, что мутация возникла среди R1 * линии в конце палеолита, возможно некоторое время между 20000 и 13000 лет назад.
Ярмарка волосы были другого физического черта связана с индоевропейцами. В отличие от этого, гены голубыми глазами уже присутствует среди мезолита европейцев, принадлежащих к Y-гаплогруппы I. Гены светлыми волосами сильнее коррелирует с распределением гаплогруппы R1a, но те, для рыжих волос не были найдены в Европе, прежде чем Бронзовый век, и, кажется, были распространяется, главным образом R1b человек (=> увидеть истоки рыжими волосами).
В материнских линий (мтДНК), соответствующие гаплогруппы R1a
Сравнивая регионы, где гаплогруппа R1a встречается сегодня с современными частот мтДНК, выясняется, что материнские клоны, которые коррелируют с большинства Y-гаплогруппы R1a являются мт-гаплогруппы C4a, H1B, H1C, H2a1, H6, H7, H11, T1a1a1, U2E, U4, U5a1a и W, а также некоторые субклады из I, J, K, T2 и V (см ниже). Древний мтДНК из Северо-Восточной Европы также подтверждает это.
Старейший мтДНК образец от предполагаемой родине R1a мужчин 30000-летний Eary современного человека от Костенки 14 месте на реке Дон на юге России, который принадлежал к гаплогруппе U2. Эта гаплогруппа встречается на низких частотах (1-2%) по всей Европе сегодня и является наиболее распространенным в России. Только два U2 субклады находятся в Европе: U2d (редкий) и U2E (наиболее распространенный). Все остальные U2 субклады обычно находятся в Южной Азии, где они могут превышать 20% населения в некоторых регионах, в частности в Пакистане и северной Индии, где R1a и R2 являются наиболее распространенным. U2 находится, насколько Юго-Восточной Азии, где Y-гаплогруппа R1a и R1b отсутствуют, но R2 присутствует. Все эти элементы в сочетании подтверждают гипотезу, что U2 был одним из первых линий базальной Y-гаплогруппы R, и, что U2 было распространено наряду R1a, R1b и R2. Обратите внимание, что 24 000-летний Mal'ta мальчик принадлежал м-гаплогруппы U и Y-гаплогруппы R *.
Гаплогруппа С также найдены среди северо-восточных европейцев, и, кажется, присутствует в регионе по крайней мере с мезолита. МтДНК C является линия обычно встречаются в Северной Азии, Восточной Азии и среди коренных американцев. В субклады из C, найденные в Европе являются C1, C4a и C5, все субклады найдены в Алтайском крае, южной Сибири и, в частности в Западной Сибири. Они могли бы быть одним из материнской происхождений Северной Азии охотников на мамонтов в позднем палеолите, наряду гаплогруппы U2. C5 был выявлен в мезолита Карелии (северо-западная Россия), в то время как C4a2 был среди линий культуры Днепровско-Донецкой в неолита Украины. Образцы C4a3 и C4a6 знакомства с бронзового века (катакомбной культуры) были также найдены в Одесской области Украины. Оба C4a и C5 распространены среди туркмен, узбеков и таджиков сегодня, населения с существенными уровнями R1a и R1b. C1 был найден на Pre-Pottery неолита сайте Расскажи Halula в северной Сирии, в регионе, где скот были впервые одомашнены. Таким образом, не исключено, что C1 принес Y-гаплогруппы R1b, которые могли бы началась в северной части Месопотамии, прежде чем перейти к понтийского степи (см историю R1b). C1 Также было обнаружено среди современных басков и каталонцев, две популяции, что полностью отсутствует Восточной Азии аутосомно примесь и Y-гаплогруппа R1a, но есть более 80% R1b. C4a и C5 более типичны для современных балто-славянских популяций, хотя их совокупный частота обычно под 1% населения. Ли и др (2010) проверили 20 мтДНК и 7 образцов Y-ДНК ранней бронзы (ок. 2000 до н.э.) остается в бассейне Тарим в Синьцзяне, на северо-западе Китая. Из этих так называемых Тарим мумий, 14 из 20 образцов мтДНК принадлежал к гаплогруппе C4, в то время как остальные шесть принадлежали M *, R * (3 проб), H и K. Все клоны Y-ДНК оказалась R1a1a.
Гаплогруппы U4 и W являются двумя клоны наиболее типичные из прибалтийско-славянских стран. Наряду с U5 и V, они, вероятно, были родные палеолитические европейские клоны, связанные с Y-ДНК гаплогруппы I, которые были ассимилированы R1a охотников на мамонтов из Северной Азии.
По мезолита, население Украины, Белоруссии и европейской части России стал бы гибрид Сибирский R1a линий мужчин и Европейский H, I, U4, U5a1, T, V и W женских линий, с лишь меньшинство сибирских материнских гаплогрупп (C4a, С5, U2). Наличие гаплогрупп С1, С4, С5, H (в том числе H17), Т, U2E, U4, U5a и U5b все были подтверждены испытаниями ДНК из мезолита европейской части России. V был найден в мезолита Иберии и Швеции. Только я и W еще не были идентифицированы в древних образцов до начала неолита. Все эти гаплогруппы сейчас можно найти в регионах, расчеты по которым индоевропейцев в эпоху бронзы и где R1a присутствует сегодня, в том числе на Кавказе, Алтае, в Средней Азии, Южной Азии и на Ближнем Востоке.
Населения Центральной Азии с повышенным процентом R1a очень полезны для перекачивания материнских линий R1a от массы других европейских гаплогрупп. Например мт-гаплогруппа J2b1a, типично европейский субклад из J, был обнаружен среди калаша из северной части Пакистана, наряду H2a1, U2E и U4. J2b1a не было найдено в другом месте к востоку от Ирана.Калаш есть 20% R1a, но нет R1b. Точно так же субклады K1b1b и K1c2 были найдены в Таджикистане, стране, которая имеет 30% R1a и только 3% R1b.
K2B был найден Кейзер соавт. (2009) в образцах бронзового века, связанных с андроновской культуры из Красноярского края на юге Сибири. Мужские образцы испытаны с того же сайта принадлежал R1a. В настоящее время K2B находится в основном в центральной и северо-восточной Европы (стран R1a) и может поэтому безопасно быть связаны с диффузии R1a ветви индоевропейцев.
H1b, H1c, H2a1 и H11, некоторые из наиболее распространенных субкладов Н в Восточной Европе и на Северном Кавказе, постоянно появляются по Сибири и Центральной Азии.
Гаплогруппа T1a1a1 является довольно распространенным в северной, центральной и восточной Европе, в первую очередь среди населения R1a. Он также находится в Иране и Пакистане, среди пуштунов, брахуи и Хунза Burusho, всех этнических групп с высоким процентом R1a, но не R1b. Вопреки T2, T1a1a1 никогда не были найдены в Европе до бронзового века, и сделать свое первое появление с культурой шнуровой керамики. Кроме T1a1a1 появились менее 7000 лет назад, едва одно тысячелетие до начала бронзового века в Понтийской степи.
Гаплогруппа T2 является более сложным, потому что его очень большим количеством субкладов. Некоторые, тем не менее может быть связано с уверенностью с бронзового века диффузии R1a. T2b2 и T2b4 были найдены в останках от культуры шнуровой керамики, и не были найдены в Европе до бронзового века. В настоящее время, как субклады встречаются в большинстве стран Европы, но T2b2 также находится на низких частотах в Иран, Туркменистан и Индия, в то время как T2b4 был найден в Азербайджане, Месопотамии, Узбекистана, Казахстана и Непала. T2b16, субклад типично северо-восточной Европе, был найден на восток до Волго-Уральского региона и Казахстана. Это, возможно, был частью расширения абашевским к Урале. T2a1b1 был найден в образцах бронзового века, связанных с андроновской культуры на юге Сибири, где образцы Y-ДНК все принадлежал R1a. Кроме того, ее современное распределение также соответствует R1a территорий, начиная от Скандинавии до Средней Азии, появляющаяся время от времени в Иране и вокруг Ближнего Востока.
Гаплогруппа W обычно встречается в регионах с высокой долей R1a и особенно часто встречается в странах Балто-славянские, в Бактрии, на севере Пакистана и северо-западной Индии. Это не было найдено в палеолита или мезолита европейских образцов. Хотя некоторые образцы W1 действительно появлялись в позднем неолите Европы (один W1 в Испании и два W1C в Германии), они могут закончили там через западным дрейфом от понтийского степи после столетий браков между соседними популяциями. Другие W субклады вдруг выскочили в Европе в эпоху бронзы, такие как в шнуровой керамики (W5A, W6a'b) и Únětice (W3a1) культур. В Индии, гаплогруппа W является значительно более распространенным среди высших каст и среди индоевропейских динамиков в соответствии с Метспалу соавт. (2004).
Субклады гаплогруппы V являются наиболее трудно отделил в тот момент, главным образом потому, много V субкладов могут быть определены лишь путем тестирования кодирующую область митохондриальной ДНК, и большинство исследований неевропейского мтДНК на сегодняшний день только проверенные Гипервариабельные область (HVR).Только субклад, которые могут быть выделены в момент, соответствующий, как R1a является V7A, который является специфическим для славянских стран и были найдены в Азербайджане
